基因合成服務(Gene Synthesis)
圖爾思技術服務中心提供的基因合成(Gene synthesis)服務,以業界最高等級的高通量oligo合成儀(1536通道)先將目標序列合成小片段oligo,然後再進行拼接,合成客戶的目標DNA序列,這種方式可以在最短的時間內產出目標DNA並大量的降低生產成本,並且能確保序列100%正確。另外,若您的目標DNA序列需要進行任何物種的序列優化,我們所使用自行開發的分析軟體能替您提供的序列進行最佳的序列優化,且圖爾思技術服務中心的序列優化服務是免費的,除此之外,我們也提供多達100多種常用的載體免費供客戶選擇,能最有效的縮短您實驗的時間,也因為我們的合成量非常大所以能提供同業所無法提供最實惠的價格。
服務特色:
1. 多達100多種免費載體可供選擇,不須額外sub-cloning (詳見載體清單)
2. 保證基因合成的序列100%準確,時間快速
3. 免費基因序列優化,使其可以在外源宿主中表達重組蛋白,以提高產量
4. 可合成難度高的DNA,包括:重複序列、高GC含量、具髮夾 結構、連續單一鹼基重複等特殊的DNA
5. 全台灣價格最低的基因合成
6. 使用新一代基因合成儀,提升30%基因合成速度
加入詢價車
服務特色:
1. 多達100多種免費載體可供選擇,不須額外sub-cloning (詳見載體清單)
2. 保證基因合成的序列100%準確,時間快速
3. 免費基因序列優化,使其可以在外源宿主中表達重組蛋白,以提高產量
4. 可合成難度高的DNA,包括:重複序列、高GC含量、具髮夾 結構、連續單一鹼基重複等特殊的DNA
5. 全台灣價格最低的基因合成
6. 使用新一代基因合成儀,提升30%基因合成速度
服務內容
基因合成是以化學合成鹼基對的方式合成重組DNA,不同於DNA和RNA合成酵素(Polymerase),基因合成並不受5'到3'合成規則的限制,可以合成5'到3'或3'到5'方向。傳統獲得DNA需要經由分子克隆(molecular cloning)和聚合酶連鎖反應(PCR)方式並由現有的DNA模板中擴增;基因合成則可以從無到有,合成一段重組DNA。
除此之外,圖爾思技術服務中心還提供完美且免費的基因序列優化服務。利用外源宿主表達重組蛋白質是現今常用的生物技術,然而其表達蛋白質的內嵌DNA可能因為包含外源宿主不常使用的密碼子,而導致重組蛋白質表現量不足。序列優化即是依照外源宿主常用的密碼子,更換重組DNA的序列,使其可以在外源宿主中表達重組蛋白,以提高產量。序列偏愛的現象主要原因為,不同物種、細胞內可用的tRNA量差異,因此選用tRNA量多的密碼子可以大幅改善重組蛋白的表達量。
客戶只需要提供欲合成的基因序列、欲表達的蛋白質序列或是NCBI上的資訊,我們將會依照您的需求幫您設計相對應的基因序列,並於最短的交付日期內提供序列100%正確無誤的產品。
除此之外,圖爾思技術服務中心還提供完美且免費的基因序列優化服務。利用外源宿主表達重組蛋白質是現今常用的生物技術,然而其表達蛋白質的內嵌DNA可能因為包含外源宿主不常使用的密碼子,而導致重組蛋白質表現量不足。序列優化即是依照外源宿主常用的密碼子,更換重組DNA的序列,使其可以在外源宿主中表達重組蛋白,以提高產量。序列偏愛的現象主要原因為,不同物種、細胞內可用的tRNA量差異,因此選用tRNA量多的密碼子可以大幅改善重組蛋白的表達量。
客戶只需要提供欲合成的基因序列、欲表達的蛋白質序列或是NCBI上的資訊,我們將會依照您的需求幫您設計相對應的基因序列,並於最短的交付日期內提供序列100%正確無誤的產品。
服務流程
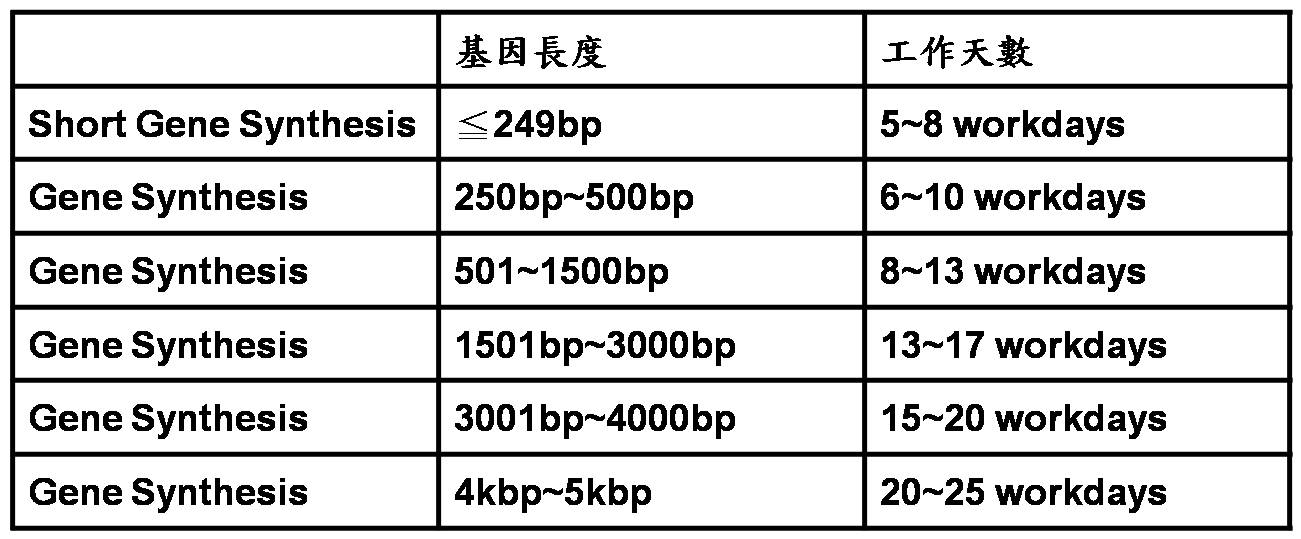
服務交期:
視合成基因之長度及結構難易度有所不同,一般約需三週,如因基因結構造成合成難度提高,合成時間會大於3週。
送件需求
1. 提供詳細且準確的DNA序列
2. 選擇5'/3' 限制酶切位
3. 確認是否要做密碼子優化 (需確認應用物種)
4. 選擇欲放入的載體
2. 選擇5'/3' 限制酶切位
3. 確認是否要做密碼子優化 (需確認應用物種)
4. 選擇欲放入的載體
出貨規格
1. 電子檔定序報告
2. 提供>5ug DNA: 含有合成基因的質體(合成產品以乾燥DNA型式交付,建議保存於-20℃)
3. 產品報告書
2. 提供>5ug DNA: 含有合成基因的質體(合成產品以乾燥DNA型式交付,建議保存於-20℃)
3. 產品報告書
免費載體列表
We offer many different kinds of vectors: the basic vector, Mammalian expression vector, E coli expression vector,Yeast expression vector, Baculovirus/Insect expression vector,Plant expression vector.....etc.
|
Cloning vector |
Mammalian expression vector |
| pUC18 | pcDNA3.0 |
| pUC19 | pcDNA3.1(+) |
| pUC57 | pcDNA3.1(-) |
| pUC57-Kan | pcDNA3.1(-)_myc-His A |
| pUC57-Simple | pcDNA3.1(-)_myc-His C |
| pUC57-Simple-NdeI-Free | pcDNA3.1(+)_myc-His A |
| pUC57-bsaI-free | pcDNA3.1(+)_myc-His B |
| PUC57-BsmbI&BsaI-free | pcDNA3.1(+)_myc-His C |
| pBlueScript II SK(+) | pcDNA3.1/Hygro(-) |
| pcDNA3.1/Hygro(+) | |
| pcDNA3.1/Zeo (+) | |
| pcDNA3.1/Zeo (-) | |
| pcDNA3.1(+)-EGFP | |
| pEGFP-C1 | |
| pEGFP-N1 | |
| pEGFP-N2 | |
| psiCHECK2.0 | |
| pGL3-Basic | |
| pGL3-Promoter | |
| pIRES2-EGFP |
|
E coli expression vector |
||
| pET-3a | pET-26b(+) | pGEX-6P-1 |
| pET-3d | pET-27b(+) | pGEX-6P-3 |
| pET-9a | pET-28a(+) | pMAL-C4X |
| pET-11a | pET-28b(+) | pMAL-C5X |
| pET-11b | pET-29a(+) | pMAL-C5E |
| pET-11d | pET-29b(+) | pMAL-p5E |
| pET-15b | PET-30a(+) | pMAl-p5X |
| pET-16b | PET-30b(+) | pACYDUET-1 |
| pET-17b | PET-31b(+) | pCDFDuet-1 |
| pET-19b | pET-32a(+) | pCOLADUET-1 |
| pET-20b(+) | pET-32b(+) | pETDuet-1 |
| pET-21a(+) | pET-41a(+) | pRSFDuet-1 |
| pET-21b(+) | pET-41b(+) | pQE-30 |
| pET-21d(+) | pET-42b(+) | pQE-60 |
| pET-22b(+) | pET-43.1a(+) | pColdI |
| pET-23a(+) | pET-45b(+) | pColdII |
| pET-24a(+) | pET-51b(+) | pBAD/His A |
| pET-24b(+) | pET-52b(+) | |
| pET-24c(+) | pGEX-4T-1 | |
| pET-24d(+) | pGEX-4T-2 | |
| pET-25b(+) | pGEX-5X-1 | |
|
Yeast expression vector |
Baculovirus/Insect |
Plant expression vector |
Gateway vector |
| pPIC3.5k | pFastBac1 | pCAMBIA1301 | pDONR207 |
| pPIC9 | pFastBac-Dual | pCAMBIA1302 | pDONR221 |
| pPIC9K | pFastBacHT A | pCAMBIA2300 | pDONR223 |
| pPICZA | pFastBacHT B | ||
| pPICZB | |||
| pPICZC | |||
| pPICZalphaA | |||
| pPICZalphaB | |||
| pPICZalphaC | |||
| pGADT7 | |||
| pGBKT7 |



