12
2022.10
蛋白質的變裝秀-轉譯後修飾(PTM)
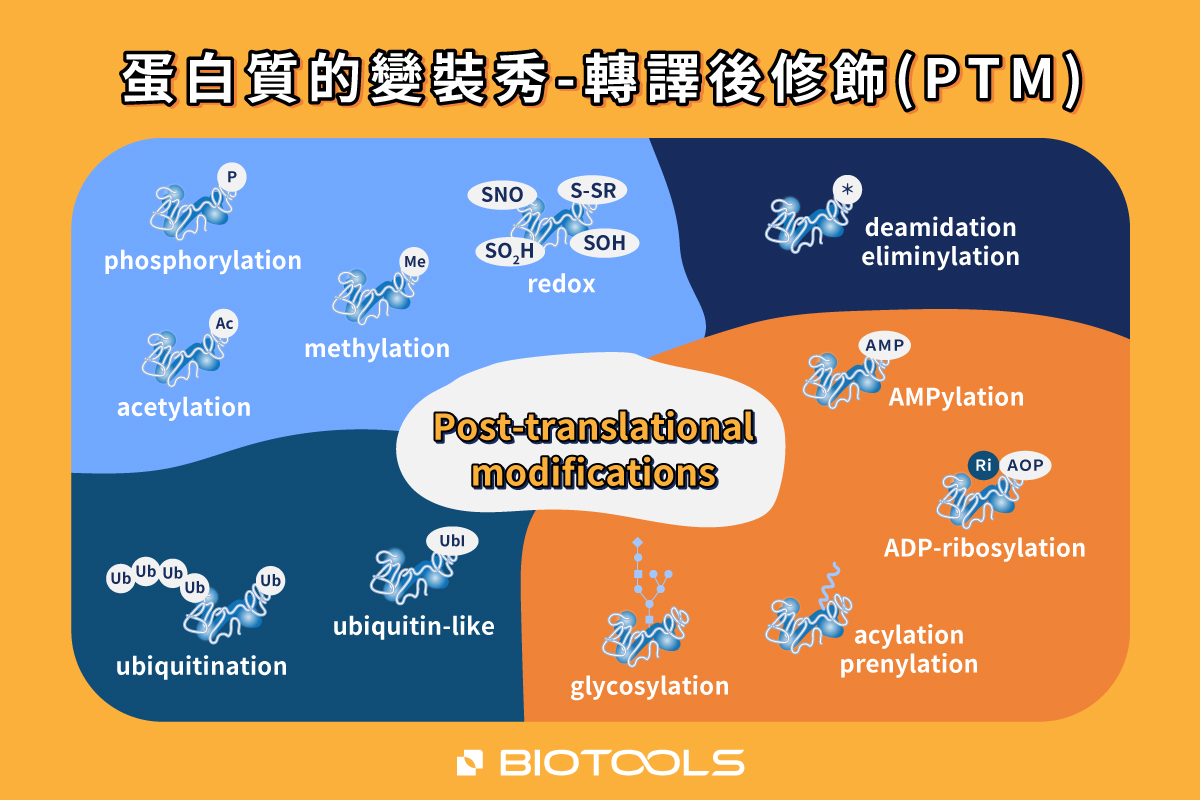
PTM會影響蛋白的行為與特徵,包含結構、功能性、壽命、交互作用、分子傳遞、受體激活、折疊與定位等,對於細胞調控扮演著相當重要的關鍵。目前已經有超過400多種不同類型的PTM,。其中最常見的PTM修飾類型有,磷酸化(phosphorylation)、甲基化(methylation)、醣基化(glycosylation)、泛素化(ubiquitination)、乙醯化(N-Acetylation)等修飾。
PTM亦和疾病息息相關,研究PTM可以幫助研究人員找到最適合的治療方式。如近年癌症研究大熱門的PD-L1,Hsu JM等研究團隊發現,PD-L1醣化會誘導癌症免疫逃避(immune invasion)與促進癌幹細胞(cancer stem cell) PD-L1的表現,透過上皮間質轉化(epithelial-mesenchymal transition, EMT) STT3的調控。另Li和Lim等研究團隊指出,非醣基化的PD-L1會ubiquitin/proteasome系統降解,因此半衰期約只有4小時,主要是因GSK3β被磷酸化所致。Lim SO等團隊證明,
CSN5抑制PD-L1的泛素化(ubiquitination)和降解(degradation),而薑黃素(curcumin)可以抑制CSN5的表達,而降低PD-L1的表現。
如何透過LC-MS/MS的檢測找到PTM修飾呢?我們可以透過蛋白質身份鑑定分析,來確認樣本中所鑑定到的蛋白種類,並透過所鑑定到的胜肽和未修飾的胜肽質量差異,找到可能的PTM與修飾位點。
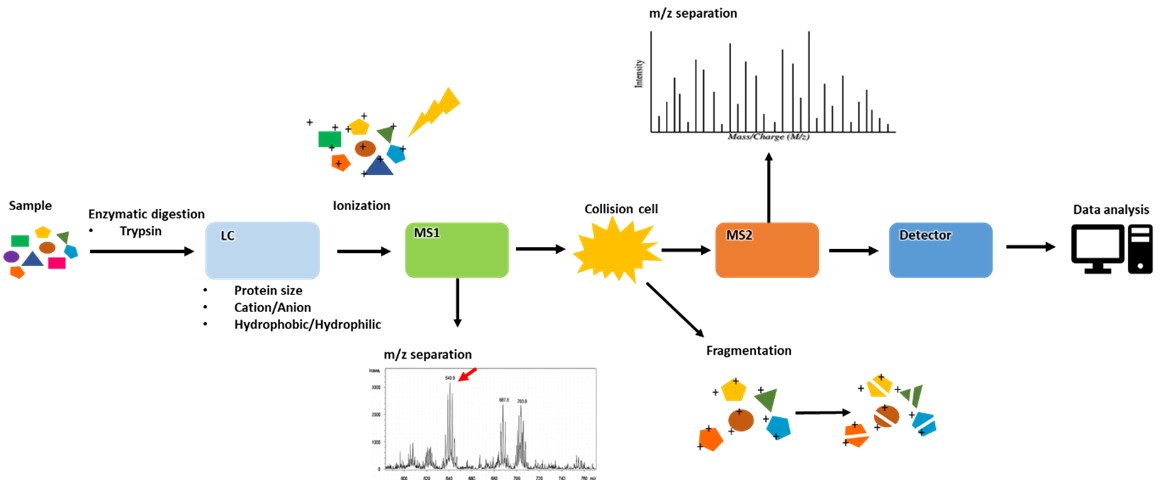 圖一、蛋白質鑑定流程
圖一、蛋白質鑑定流程
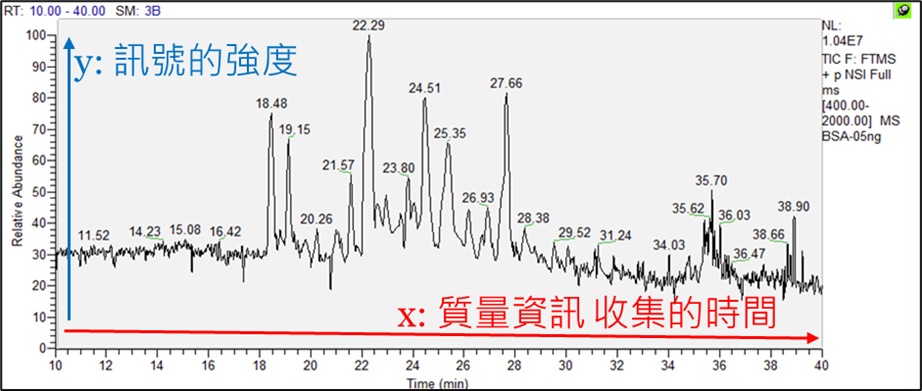 圖二、BSA層析圖
圖二、BSA層析圖
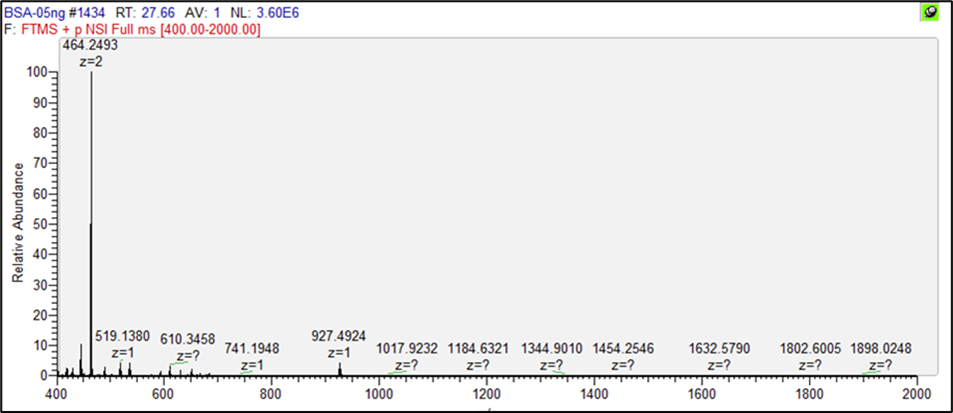 圖三、一級質譜圖(MS1)
圖三、一級質譜圖(MS1)
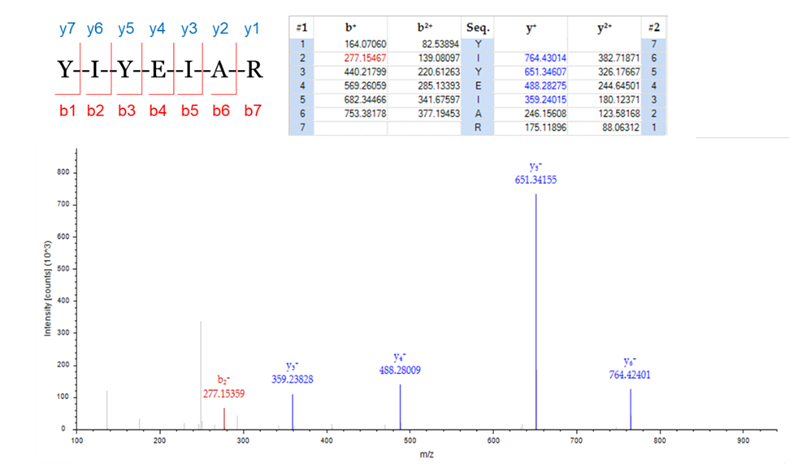 圖四、二級質譜圖(MS2)
圖四、二級質譜圖(MS2)
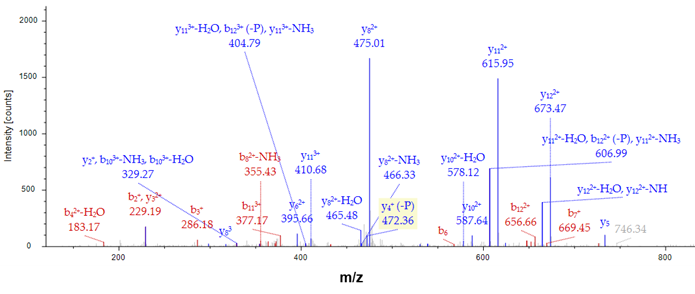
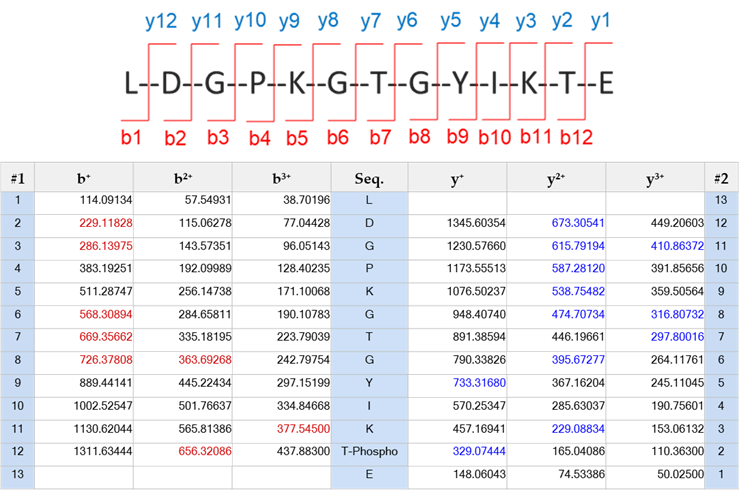 圖五、觀察磷酸化胜肽圖譜與數值
圖五、觀察磷酸化胜肽圖譜與數值
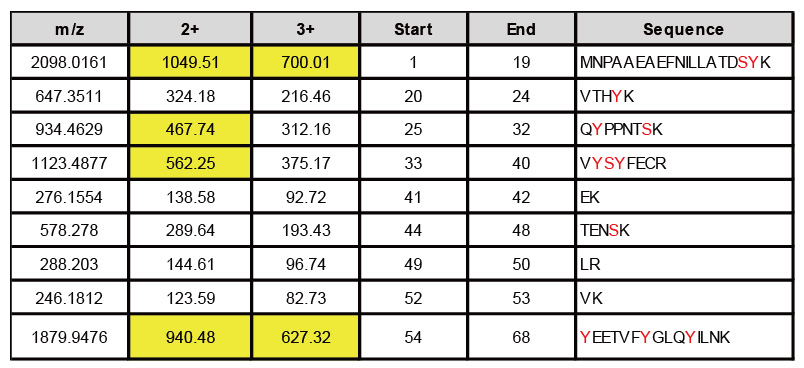 表一、蛋白經Trypsin水解預切位分析
表一、蛋白經Trypsin水解預切位分析
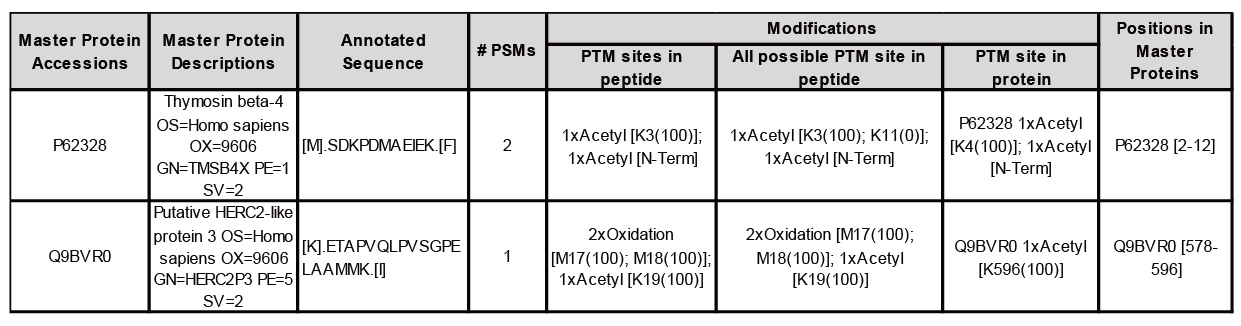 表二、乙醯化鑑定結果(參考acetylation peptide report)
表二、乙醯化鑑定結果(參考acetylation peptide report)
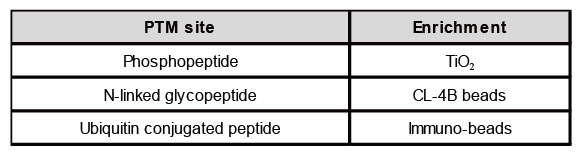 表三、PTM enrichment
表三、PTM enrichment
Reference:
PTM亦和疾病息息相關,研究PTM可以幫助研究人員找到最適合的治療方式。如近年癌症研究大熱門的PD-L1,Hsu JM等研究團隊發現,PD-L1醣化會誘導癌症免疫逃避(immune invasion)與促進癌幹細胞(cancer stem cell) PD-L1的表現,透過上皮間質轉化(epithelial-mesenchymal transition, EMT) STT3的調控。另Li和Lim等研究團隊指出,非醣基化的PD-L1會ubiquitin/proteasome系統降解,因此半衰期約只有4小時,主要是因GSK3β被磷酸化所致。Lim SO等團隊證明,
CSN5抑制PD-L1的泛素化(ubiquitination)和降解(degradation),而薑黃素(curcumin)可以抑制CSN5的表達,而降低PD-L1的表現。
如何透過LC-MS/MS的檢測找到PTM修飾呢?我們可以透過蛋白質身份鑑定分析,來確認樣本中所鑑定到的蛋白種類,並透過所鑑定到的胜肽和未修飾的胜肽質量差異,找到可能的PTM與修飾位點。
蛋白質身分鑑定
蛋白質鑑定分析會怎麼進行呢?首先我們會先將樣本經過Trypsin水解,得到小片段的胜肽(peptide),每個蛋白都會有獨特的序列(unique peptide),而可透過質譜圖中的胜肽質量分佈,以資料庫(database)和搜尋引擎(Mascot)進行胜肽質量比對,並以統計方式(bottom-up protein assembly)找出最有可能的蛋白序列。因此透過這樣的方式可以先找到樣本中的目標蛋白,才可以深入探討有可能的PTM位點。圖一為蛋白質鑑定的流程。
 圖一、蛋白質鑑定流程
圖一、蛋白質鑑定流程處理好的樣本會透過HPLC將複雜的樣本先以極性與電荷,將胜肽進行初步的分層如圖二,可獲得層析圖(chromatogram),其主要是以收集的時間(Retention time)來蒐集樣本中質量的資訊。
 圖二、BSA層析圖
圖二、BSA層析圖而經管柱流出的胜肽,會透過電噴灑噴灑游離法(ESI),將胜肽帶電,進入到質譜儀偵測,因此可以獲得一級質譜圖(MS1)如圖三,在27.6分鐘所記錄到的胜肽質量(MS)有464.2493、927.4924等等,其中BSA樣本當中927.4924是在27.6分鐘當中,記錄到最大的胜肽質量,而464.2493為訊號最強。
 圖三、一級質譜圖(MS1)
圖三、一級質譜圖(MS1)接著胜肽會透過更高能量(CID)打碎裂成更小不同系列的離子,y系列離子斷裂在不同位置氨端碎片(N-terminal fragment),b系列離子則為羧端碎片(C-terminal fragment),數字則代表胺基酸的排列順序,並透過y1、y2等的質量差距可以計算出該胜肽的序列。而游離出來的離子帶多電荷,如一價、二價、三價等,一般來說帶二價以上的離子產生的二級質譜(MS/MS, MS2)品質較佳。下圖四為一個7個胺基酸的胜肽,b2離子就是YI、y5就是YEIAR,因此b2+y5會組成這段完整的胜肽序列(YIYEIAR)。
 圖四、二級質譜圖(MS2)
圖四、二級質譜圖(MS2)PTM的鑑定與分析
今天我們可以從圖五看到樣本中的實際值(右圖MS/MS圖譜)和理論值(左表)經過比對(PSM,peptide-spectrum match)後的結果,可以鑑定到的碎片如紅色(b系列離子)與藍色(y系列離子)的標示,而黑色標示則為理論值,故經分析比對該胜肽序列為LDGPKGTGYIKTE,同時也可以從圖譜中觀察到磷酸化的位點,其y2離子有被鑑定到,因此在T的位點上是有磷酸化的修飾。

 圖五、觀察磷酸化胜肽圖譜與數值
圖五、觀察磷酸化胜肽圖譜與數值酵素預切位判定
若想了解某個蛋白的磷酸化位點(S、Y、T),可以先進行酵素預切位的分析,而目前最常使用的酵素為Trypsin,主要可以剪切的位置為Lysine (K)與Arginine (R),而序列偵測的範圍會依照每台儀器的靈敏度而有所不同。以Oribitrap Elite為例,當我們進行酵素預切位分析(表一),在被鑑定到的序列中以黃底標示的為可被鑑定到的序列,而其他序列可能會因片段過小或過大,超出儀器偵測範圍,因此我們感興趣的位點是Y26則可以被鑑定到,但是位點在S47則無法被鑑定到。
 表一、蛋白經Trypsin水解預切位分析
表一、蛋白經Trypsin水解預切位分析
若無法透過Trypsin找到目標的位點,或超出儀器可以偵測的範圍,可以建議更換成其他的酵素,如Glu-c剪切位置為Glutamate (E)、Lys-C剪切位置為Lysine (K)、Asp-N剪切位置為Aspartate (D)等進行預測。
PTM分析與報告判讀
在圖爾思蛋白質體學服務當中,會根據客戶的樣本進行樣本前處理,再進行上樣與分析,而目前有磷酸化、醣化、泛素化、甲基化等的PTM修飾分析,並除了目標蛋白以外,樣本中所鑑定到的蛋白,可能的PTM修飾位點。如表二範例所示,我們可以觀察到蛋白Thymosin beta-4中,可在SDKPDMAEIEK的胜肽中,鑑定到K3位點上有乙醯化(acetylation),且被鑑定到的機率有100%,K11位點上有乙醯化機率為0%,因此可以確定K3是有乙醯化的修飾;而在蛋白HERC2P3中,可在ETAPVQLPVSGPELAAMMK的胜肽中,鑑定到M17、M18、K19有乙醯化的機率也有100%。
PTM分析與報告判讀
在圖爾思蛋白質體學服務當中,會根據客戶的樣本進行樣本前處理,再進行上樣與分析,而目前有磷酸化、醣化、泛素化、甲基化等的PTM修飾分析,並除了目標蛋白以外,樣本中所鑑定到的蛋白,可能的PTM修飾位點。如表二範例所示,我們可以觀察到蛋白Thymosin beta-4中,可在SDKPDMAEIEK的胜肽中,鑑定到K3位點上有乙醯化(acetylation),且被鑑定到的機率有100%,K11位點上有乙醯化機率為0%,因此可以確定K3是有乙醯化的修飾;而在蛋白HERC2P3中,可在ETAPVQLPVSGPELAAMMK的胜肽中,鑑定到M17、M18、K19有乙醯化的機率也有100%。
 表二、乙醯化鑑定結果(參考acetylation peptide report)
表二、乙醯化鑑定結果(參考acetylation peptide report)此外目前對磷酸化(phos-site)、醣化(N-linked Glyco-site)與泛素化(Ubiquitin-site)位點分析,會經過富集(enrichment)步驟處理,來增強蛋白鑑定與富集的能力,使蛋白的分辨率更高,從而獲得更精確的PTM結果,如表三。
 表三、PTM enrichment
表三、PTM enrichment
還在煩惱怎麼證明PTM的存在呢?小編大力推薦圖爾思提供的技術服務「蛋白質體學服務」,已經有超過700多個服務案件的經驗,並擁有鑑定超過900多種PTM修飾的服務,以及優良的售前與售後專業服務,解決你所有的疑難雜症,產出優良的實驗數據!還在等什麼點擊以下網址!!
Reference:
- Ramazi S, Zahiri J. Posttranslational modifications in proteins: resources, tools and prediction methods. Database (Oxford). 2021 Apr 7;2021:baab012. doi: 10.1093/database/baab012. PMID: 33826699; PMCID: PMC8040245.
- Hsu JM, Xia W, Hsu YH, Chan LC, Yu WH, Cha JH, Chen CT, Liao HW, Kuo CW, Khoo KH, Hsu JL, Li CW, Lim SO, Chang SS, Chen YC, Ren GX, Hung MC. STT3-dependent PD-L1 accumulation on cancer stem cells promotes immune evasion. Nat Commun. 2018 May 15;9(1):1908. doi: 10.1038/s41467-018-04313-6. PMID: 29765039; PMCID: PMC5954021.
- Li CW, Lim SO, Xia W, Lee HH, Chan LC, Kuo CW, Khoo KH, Chang SS, Cha JH, Kim T, Hsu JL, Wu Y, Hsu JM, Yamaguchi H, Ding Q, Wang Y, Yao J, Lee CC, Wu HJ, Sahin AA, Allison JP, Yu D, Hortobagyi GN, Hung MC. Glycosylation and stabilization of programmed death ligand-1 suppresses T-cell activity. Nat Commun. 2016 Aug 30;7:12632. doi: 10.1038/ncomms12632. PMID: 27572267; PMCID: PMC5013604.
- Lim SO, Li CW, Xia W, Cha JH, Chan LC, Wu Y, Chang SS, Lin WC, Hsu JM, Hsu YH, Kim T, Chang WC, Hsu JL, Yamaguchi H, Ding Q, Wang Y, Yang Y, Chen CH, Sahin AA, Yu D, Hortobagyi GN, Hung MC. Deubiquitination and Stabilization of PD-L1 by CSN5. Cancer Cell. 2016 Dec 12;30(6):925-939. doi: 10.1016/j.ccell.2016.10.010. Epub 2016 Nov 17. PMID: 27866850; PMCID: PMC5171205.



